Transandinomys talamancae - Transandinomys talamancae
| Transandinomys talamancae | |
|---|---|
 | |
| Череп мужчины из Гатуна, Панама, вид сверху[1] | |
| Научная классификация | |
| Королевство: | Animalia |
| Тип: | Хордовые |
| Учебный класс: | Млекопитающие |
| Заказ: | Rodentia |
| Семья: | Cricetidae |
| Подсемейство: | Sigmodontinae |
| Род: | Трансандиномис |
| Разновидность: | T. talamancae |
| Биномиальное имя | |
| Transandinomys talamancae (J.A. Аллен, 1891) | |
 | |
| Распределение Transandinomys talamancae в южной части Центральной Америки и северо-западе Южной Америки[3] | |
| Синонимы[13] | |
| |

Transandinomys talamancae это грызун в род Трансандиномис что происходит из Коста-Рика на юго-запад Эквадор и северный Венесуэла. Его среда обитания состоит из низинных лесов на высоте до 1500 м (5000 футов) над уровнем моря. уровень моря. С телом масса от 38 до 74 г (от 1,3 до 2,6 унций), это средний рисовая крыса. Мех мягкий, от красноватого до коричневатого на верхней стороне и от белого до белого. бафф на нижние части. В хвост сверху темно-коричневый, снизу светлее, уши и лапы длинные. В вибриссы (усы) очень длинные. В черепе длинная головотрубка (передняя часть) и черепная коробка низкий. Количество хромосомы варьируется от 34 до 54.
Впервые вид был описан в 1891 г. Джоэл Асаф Аллен и после этого множество имен, теперь считающихся синонимы, были применены к местному населению. Это было сосредоточенный в широко распространенный вид »Oryzomys capito" (сейчас же Hylaeamys megacephalus ) с 1960-х по 1980-е годы, а нынешнее распределение синонимов датируется 1998 годом. Он был помещен в род Оризомыс до 2006 года, как Oryzomys talamancae, но не имеет непосредственного отношения к типовой вид этого рода и поэтому был переведен в отдельный род Трансандиномис в 2006 году. Разделяет этот род с Transandinomys bolivaris с еще более длинными вибриссами; два пересекаются в широком смысле в распределении и являются морфологически похожий.
Активен ночью, Transandinomys talamancae живет на земле и питается растениями и насекомыми. Самцы больше двигаются и имеют больше домашние диапазоны чем у большинства женщин. Гнездится круглый год, хотя немногие особи выживают больше года. После беременность В период около 28 дней рождаются от двух до пяти детенышей, которые достигают половой зрелости в течение двух месяцев. Разнообразие паразиты встречаются на этом виде. Широко распространен и обычен, не представляет угрозы для сохранения.
Таксономия
В 1891 г. Джоэл Асаф Аллен был первым, кто научно описал Transandinomys talamancae, когда он назвал Oryzomys talamancae из экземпляра из Таламанки, Коста-Рика. Он поместил его в род Оризомыс, тогда более широко, чем сейчас, и сравнил его с обоими болотная рисовая крыса (О. palustris) и к О. laticeps.[14] Несколько других имен, которые теперь известны как синонимы из Transandinomys talamancae были введены в последующие годы. В 1899 году Аллен описал Oryzomys mollipilosus, О. magdalenae, и О. villosus из Магдалена, Колумбия.[15] Олдфилд Томас добавлен О. sylvaticus из Санта-Роза, Эквадор в 1900 г.[16] и О. panamensis из Панама, Панама, 1901 год.[17] В том же году Вирт Робинсон и Маркус Лайон назвали Oryzomys medius с близкого расстояния Ла Гуайра, Венесуэла.[18] Аллен добавил О. каррикери из Таламанки, Коста-Рика, в 1908 году.[19]
Эдвард Альфонсо Гольдман пересмотренный североамериканский Оризомыс в 1918 г. Он поместил оба Panamensis и Carrikeri как синонимы Oryzomys talamancae и упомянул О. mollipilosus и О. medius как близкородственные виды. О. таламанка был единственным представителем своей собственной видовой группы, которую Голдман считал наиболее близкой к Oryzomys bombycinus (=Transandinomys bolivaris ).[20] В 1960 г. Филип Гершковиц перечисленные Talamancae, средний, Magdalenae, sylvaticus, и mollipilosus среди множества синонимов слова "Оризомыс латицепс", имя позже заменено на"Oryzomys capito".[21] Вид оставался сосредоточенным в Oryzomys capito до 1983 года, когда Альфред Гарднер снова перечислил его как допустимый вид, действие более полно задокументировано Гай Мюссер и Марина Уильямс в 1985 году.[22] Мюссер и Уильямс также обнаружили, что голотип из Oryzomys villosus, родство которых оспаривалось, на самом деле состояло из кожи О. таламанка и череп Oryzomys albigularis группа (эквивалент текущего рода Нефеломис ). Они ограничили название скином, сделав виллоз синоним О. таламанка.[23] Они также исследовали голотипы Panamensis, Carrikeri, mollipilosus, средний, и Magdalenae и назвал их примерами Oryzomys talamancae. Кроме того, они включали сильватический и Oryzomys castaneus J.A. Allen, 1901, из Эквадора как синонимы, но без изучения голотипов.[24] Мюссер и его коллеги повторно рассмотрели группу в 1998 году и подтвердили, что сильватический представляет Oryzomys talamancae; однако они обнаружили, что Castaneus на самом деле был примером Оризомыс боливарис (электрический ток Transandinomys bolivaris).[25]
В 2006 году Марсело Векслер опубликовал филогенетический анализ Oryzomyini («рисовые крысы»), племя которому Оризомыс выделяется, используя морфологический данные и Последовательности ДНК от IRBP ген. Его результаты показали виды Оризомыс рассредоточены по Oryzomyini и предположили, что большинство видов этого рода следует отнести к новым родам.[26] Oryzomys talamancae также был включен; он появился в "клады B "вместе с другими видами, ранее связанными с Oryzomys capito. Некоторые анализы поместили его ближе всего к видам, которые сейчас помещаются в Эвриоризомыс или же Нефеломис, но с низкой поддержкой.[27] Позже в том же году он вместе с Александром Персекильо и Робертом Воссом назвал десять новых родов видов, ранее помещенных в Оризомыс, включая Трансандиномис, у которого есть Oryzomys talamancae (сейчас же Transandinomys talamancae) как его типовой вид.[12] Они также включали Оризомыс боливарис, который не был включен в филогенетическое исследование Векслера, в этот новый род. Эти два вида морфологически похожи, но Векслер и его коллеги смогли идентифицировать только один. синапоморфия (общая-производная черта) для них: очень долго надбровные вибриссы (вибриссы, или усы над глазами).[28] Трансандиномис является одним из примерно 30 родов Oryzomyini, разнообразного сообщества американских грызунов, насчитывающего более ста видов,[29] и на более высоких таксономических уровнях в подсемействе Sigmodontinae семьи Cricetidae, наряду с сотнями других видов, в основном мелких грызунов.[30]
Несколько общие имена были предложены для Transandinomys talamancae, в том числе "Рисовая крыса Таламанка",[31] "Трансандийский Оризомыс",[32] и «Таламанканская рисовая крыса».[2]
Описание
Transandinomys talamancae это ярко окрашенная рисовая крыса среднего размера.[33] Это похоже на Т. bolivaris и их часто путают.[34] Они примерно такие же большие, но в T. talamancae хвост длиннее, а задние лапы короче.[35] У обоих видов есть уникальные длинные вибриссы, при этом как мистициальные (над ртом), так и надбровные вибриссы простираются до или за задний край ушей, когда они лежат спиной к голове, но Т. bolivaris существенно длиннее.[12] Х. альфарои, широко распространенный вид от Мексики до Эквадора, также похож.[36] Он меньше и темнее, но молодой взрослый T. talamancae похожи по цвету на взрослых Х. альфарои и часто ошибочно идентифицируются.[37] Hylaeamys megacephalus,[Примечание 1] с которым T. talamancae был синонимом в течение нескольких десятилетий, похож по размеру тела, но, как известно, не пересекается с T. talamancae в диапазоне.[39]
Мех короткий, густой и мягкий в Transandinomys talamancae;[40] в Т. bolivaris, он длиннее и еще мягче и плотнее.[41] Цвет верхней части тела варьируется от красноватого до коричневатого, становясь светлее к бокам и щекам.[31] Нижняя сторона белая до бафф, с основаниями волосков пухлый (свинцового цвета).[31] Мех Т. bolivaris темнее: сверху темно-коричневый, снизу темно-серый.[42] H. megacephalus также имеет более темный мех.[43] У молодых особей тонкий серый мех, который линял в темно-коричневый мех подростков, когда животному от 35 до 40 дней. Этот мех заменяется ярким мехом взрослых в возрасте от 49 до 56 дней.[44] Молодь никогда не бывает черноватой, как в Т. bolivaris.[42] Уши темно-коричневые,[40] большие, густо покрытые очень мелкими волосками.[45]
| Страна | п[Заметка 2] | Голова и тело | Хвост | Задняя лапа | Ухо |
|---|---|---|---|---|---|
| Панама | 22 | 124.4 (101–136) | 124.7 (110–143) | 29.2 (27–32) | 21.1 (16–25) |
| Колумбия | 13 | 135.2 (120–151) | 125.2 (114–140) | 29.5 (27–32) | 19.0 (18–22) |
| Эквадор | 20[Заметка 3] | 124 (118–136) | 128.5 (118–137) | 29.1 (28–31) | – |
| Размеры даны в миллиметрах и в форме «среднее (минимум – максимум)». | |||||
Редко опушенный хвост примерно такой же длины, как голова и тело.[45] Сверху он темно-коричневый, а снизу светлее.[40] Напротив, хвост H. megacephalus практически не имеет разницы в цвете между верхней и нижней поверхностью.[43] В 2006 году Векслер и его коллеги отметили окраску хвоста как разницу между двумя видами Трансандиномис (двухцветный в T. talamancae и одноцветный в Т. bolivaris),[12] но в своем исследовании 1998 года Мюссер и его коллеги не смогли найти различий в окраске хвоста между панамскими образцами этих двух видов.[47]
Задние лапы длинные, у них три центральных пальца длиннее двух внешних.[47] Сверху они от белого до бледно-желтого,[40] где ступня покрыта волосами, которые длиннее, чем у Т. bolivaris.[48] Пальцы задних лап окружены ногтевые пучки серебристых волос, длиннее самих когтей.[49] Когти короткие и острые.[50] Части подошвы покрыты нечеткими чешуйками (чешуйки ), которые обычно полностью отсутствуют в Т. bolivaris.[51] В колодки умеренно большие.[47]
Длина головы и тела от 105 до 151 мм (от 4,1 до 5,9 дюйма), длина хвоста от 105 до 152 мм (от 4,1 до 6,0 дюйма), длина задней части стопы от 26 до 32 мм (от 1,0 до 1,3 дюйма), длина ушей от 17 до 24 мм (от 0,67 до 0,94 дюйма), а масса тела от 38 до 74 г (от 1,3 до 2,6 унции).[52] Как и у большинства оризомиинов, у самок восемь молочные железы.[53] Всего 12 грудной позвонок со связанными ребрами, 7 поясницы, и 29 хвостовой; изредка присутствует пара лишних (дополнительных) ребер.[54]
Череп и зубы
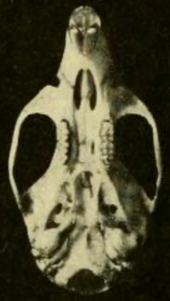
Череп имеет длинный рострум (переднюю часть), широкий межглазничная область (между глазами) и низкий черепная коробка.[55] Он отличается от Т. bolivaris в различных пропорциях, которые более заметны у взрослых, чем у молодых;[48] череп H. megacephalus заметно больше.[43] В скуловая пластина широкая, с хорошо развитой скуловой выемкой спереди. Его задний край находится на уровне передней части первой верхней коренной зуб.[56] В скуловая дуга (скула) тяжелая. В носовой и предчелюстной кости отходят примерно так же далеко назад.[31] Межглазничная область наиболее узкая к передней части и имеет слабые бусинки по краям;[57] Т. bolivaris похожа, но имеет более сильную отбортовку[58] и H. megacephalus полностью отсутствует бисероплетение.[59] В теменная кость обычно ограничивается крышей черепной коробки и не простирается вбок, как в большинстве Т. bolivaris.[58] В меж теменная кость, часть крыши черепной коробки, большая.[31]
В резкое отверстие (проемы в передней части нёбо ) короткие и не доходят до первых моляров;[40] они дольше в Х. альфарои.[37] Костное небо длинное и выходит за пределы конца ряда коренных зубов и заднего края моляров. верхнечелюстной кости.[60] В заднебоковые небные ямки, которые перфорируют нёбо около третьих моляров, имеют небольшой размер и могут углубляться или не углубляться в ямка.[61] В клиновидно-небные пустоты (проемы в крыше мезоптеригоидная ямка, за нёбом) тоже мелкие,[31] как и слуховые буллы.[40] Как и у большинства оризомиинов, подсквамозное отверстие, отверстие в задней части черепа.[62] Рисунок пазов и отверстие (отверстия) в черепе указывает на то, что кровообращение артерий головы в T. talamancae следует примитивному образцу, как и у большинства подобных видов, но в отличие от Hylaeamys.[59]
В нижняя челюсть (нижняя челюсть) менее прочная, чем в Т. bolivaris.[63] В венечный отросток (а процесс в задней части кости) небольшая, а капсульный отросток, в котором находится корень нижнего резец, маленькие.[31] В подбородочное отверстие, расположенный в диастема между нижним резец и первый моляр открывается в сторону, как обычно у оризомиинов.[64] Верхний и нижний жевательные гребни, которые закрепляют некоторые из жевательных мышц, не соединяются в один гребень и простираются вперед ниже первого моляра.[65]
Верхний резец опистодонт, с режущей кромкой, направленной назад.[66] Как обычно у oryzomyines, коренные зубы Brachydont (с низкой короной) и бунодонт (с выступами выше соединительных гребней).[67] Первый верхний моляр уже, чем у Т. bolivaris. У этого вида, но в отличие от многих других рисовых крыс, в том числе Х. альфарои и E. nitidus, то мезофлексия на втором верхнем моляре, который разделяет паракон (один из главных куспидов) от мезолоф (вспомогательный гребень), не разделяется эмалевым мостом надвое.[63] В гипофлексид на втором нижнем моляре основная впадина между бугорками очень длинная, простирается более чем на половину длины зуба; по этому признаку вид снова похож на Т. bolivaris но в отличие от Х. альфарои.[68] Каждый из верхних коренных зубов имеет три корня (два на губной или внешней стороне и один на язычной или внутренней стороне), а у каждого из нижних коренных зубов - по два (один спереди и один сзади); T. talamancae не имеет дополнительных мелких корней, которые присутствуют у различных других оризомиинов, включая виды Эвриоризомыс, Нефеломис, и Handleyomys.[69]
Анатомия мужской репродуктивной системы

Как характерно для Sigmodontinae, Transandinomys talamancae имеет сложный половой член, с дистальным (дальним) концом бакулюм (кость пениса) оканчивается структурой, состоящей из трех пальцев.[71] Как и у большинства оризомиинов, центральный палец больше двух по бокам.[72] Наружная поверхность полового члена в основном покрыты мелкими шипами, но есть широкая полоса немостистой ткани.[73]
Некоторые особенности добавочных желез в области мужских половых органов у оризомиинов различаются. В Transandinomys talamancae,[Примечание 4] одна пара препуциальные железы присутствует на половом члене. Как обычно у сигмодонтин, есть две пары вентральный предстательная железа железы и одна пара передний и спинной предстательные железы. Часть конца везикулярная железа сложен неправильно, не гладкий, как у большинства оризомиинов.[75]
Кариотип

В кариотип в T. talamancae переменная. Образцы из двух разных мест в Венесуэле содержат 34хромосомы и основное число 64 хромосомных плеч (2n = 34, FN = 64).[77] Каждый из четырех экземпляров из другого населенного пункта Венесуэлы имеет свой кариотип с числом хромосом от 40 до 42 и основным числом от 66 до, возможно, 67.[Примечание 5] В аутосомы (неполовые хромосомы ) кариотипа 2n = 34 имеют два основных плеча, но кариотипы 2n = 40–42 включают несколько акроцентрический аутосомы, у которых есть только одна большая рука. Кариотип 2n = 34 включает два больших субметацентрический пары, у которых две длинные руки, но одна заметно длиннее другой, и одна пара субтелоцентрический хромосомы с длинным и намного более коротким плечом, но кариотипы 2n = 40–42 лишены субметацентриков и имеют другую пару субтелоцентриков.[79] Обе Робертсоновские транслокации (слияние длинного плеча одной хромосомы с длинным плечом другой и короткого плеча одной хромосомы с коротким плечом другой) и перицентрические инверсии (инверсия части хромосомы, которая включает центромера ) необходимы, чтобы объяснить разницу между двумя группами. Musser и его коллеги, обсуждая эти кариотипы, предположили, что образец 2n = 40–42 был изнутри гибридная зона между двумя кариотипическими морфами.[80]
Кариотип образца Эквадора с севера Залив Гуаякиль аналогичен венесуэльским животным при 2n = 36, FN = 60; он включает четыре акроцентрических и две субтелоцентрических пары и никаких субметацентрических пар. Напротив, образец с юга залива имел 2n = 54, FN = 60, включая 23 пары акроцентрики и четыре пары акроцентрики. метацентрики (с двумя одинаково длинными руками). Мюссер и его коллеги назвали разницу между двумя эквадорскими формами «впечатляющей».[80] и отметил, что необходимы дальнейшие исследования для более полного понимания кариотипической дифференциации внутри вида.[81] Обе Т. bolivaris и Х. альфарои имеют больше хромосом и плеч, при 2n = 58, FN = 80 и 2n = 60–62, FN = 100–104 соответственно.[82] Hylaeamys megacephalus имеет 2n = 54, FN = 58–62 и аналогичные Hylaeamys perenensis имеет 2n = 52, FN = 62; эти кариотипы напоминают кариотипы южных эквадорских T. talamancae.[83]
Распространение и среда обитания
Распределение Transandinomys talamancae простирается от северо-запада Коста-Рики на юг и восток до севера Венесуэлы и юго-запада Эквадора, до 1500 м (5000 футов) над уровнем моря.[32] Это лесной вид, встречается как в вечнозеленый и лиственный лес.[84] Хотя его распространение во многом перекрывает распространение Т. bolivaris, он более широко распространен в Южной Америке из-за большей устойчивости к средам обитания в сухих лесах.[85]
Transandinomys talamancae достигает северной границы своего ареала в Коста-Рике, но за исключением одной записи с крайнего северо-запада (в Провинция Гуанакасте около южной окраины Озеро Никарагуа ), он известен только с юго-востока страны. В отличие, Т. bolivaris и Х. альфарои происходят дальше на север, в Гондурас и Мексику соответственно. Это происходит по всей Панаме на небольших возвышенностях. Вдоль тихоокеанского побережья в Колумбии и Эквадоре он встречается на прибрежной равнине и прилегающих предгорьях Анд.[85] Самая южная известная находка находится на далеком юго-западе Эквадора, но этот вид может распространяться и в соседнее Перу.[84]
Он также встречается по всей северной Колумбии на низких высотах и в западной Венесуэле к западу от Озеро Маракайбо и у подножия западной части Венесуэльский прибрежный хребет на восток до Национальный парк Гуатопо.[85] Hylaeamys megacephalus встречается дальше на восток в восточной части прибрежного хребта, отделенного прибрежным Восточно-карибская засушливая зона.[43] Есть запись из Дельта Ориноко северо-востока Венесуэлы, вполне в пределах Hylaeamys megacephalus, но Массер и его коллеги предполагают, что это основано на неправильно маркированных образцах.[86] Также вид был обнаружен на узкой полосе между Льянос и Анды (Кордильеры Восточные и Кордильера-де-Мерида ) в восточной Колумбии и на северо-западе Венесуэлы.[85] Безлесный Льянос отделяет эти районы от Hylaeamys населения. Hylaeamys perenensis[Примечание 6] однако встречается южнее, вдоль восточных предгорий Восточных Кордильер в Колумбии, и возможно, что они пересекаются в этой области.[43]
Экология и поведение
Transandinomys talamancae - обычный, даже многочисленный вид.[88] Его экологию изучал Теодор Флеминг в Зона Панамского канала.[89] Он живет на земле и активен ночью.[90] Гнездо животных находится выше уровня земли и иногда попадает в норы, которые также используются карманной мышью. Liomys adspersus.[91] Его диета всеядный: включая растительный материал, такой как семена и фрукты; взрослые и личинки насекомых.[44]
Самцы обычно преодолевают большие расстояния, чем самки. Средний размер жилого квартала в исследовании Флеминга составлял 1,33.гектаров (3.3 акры ); у самцов в среднем были большие домашние ареалы.[92] Однажды отловленные экземпляры, как правило, отлавливаются чаще, чем никогда не отловленные.[93] Флеминг подсчитал, что плотность населения достигла пика до 4,3 на га (1,7 на акр) в конце сезона дождей (октябрь – ноябрь), но упала почти до нуля примерно в июне; однако эти цифры вполне могут быть заниженными.[94] В центральной Венесуэле плотность населения колеблется от 5,5 до 9,6 на га (от 2,2 до 3,8 на акр).[95]
В Панаме этот вид размножается круглый год без явной сезонной изменчивости.[96] По словам Омара Линареса Mamíferos de Venezuela (Млекопитающие Венесуэлы), наибольшая репродуктивная активность приходится на июнь – июль и декабрь.[95] В лаборатории период беременности составляет 28 дней;[97] Линарес сообщает, что в дикой природе от 20 до 30 дней.[95] Самки производят в среднем шесть пометов в год и от двух до пяти (в среднем 3,92) детенышей в помете, так что одна самка может произвести около 24 детенышей в год;[98] это, вероятно, завышенная оценка, поскольку большинство самок не проживут целый год. Более крупные самки могут иметь больший помет.[99] Животные становятся половозрелыми в возрасте менее двух месяцев; Согласно исследованию Флеминга, некоторые самки в молодом мехе, вероятно, младше 50 дней, уже были беременны.[100] Самому старому экземпляру, который наблюдал Флеминг, было девять месяцев;[101] он подсчитал, что животные вряд ли проживут более года в дикой природе[102] и что средний возраст смерти составлял 2,9 месяца.[103]
Десять видов клещей (Gigantolaelaps aitkeni,[104][Примечание 7] Gigantolaelaps gilmorei, Gigantolaelaps oudemansi, Gigantolaelaps wolffsohni, Haemolaelaps glasgowi, Лаэлапс деармаси, Лаэлапс пилифер, Лаэлапс тори, Mysolaelaps parvispinosus,[110] и Paraspeleognathopsis cricetidarum ),[111] тринадцать чиггеров (Aitkenius cunctatus,[112] Аскошоэнгастия дискрита, Eutrombicula alfreddugesi, Eutrombicula goeldii, Intercutestrix tryssa, Leptotrombidium panamensis, Миксакарус осциллатус, Pseudoschoengastia abditiva, Pseudoschoengastia bulbifera, Trombicula dunni, и Trombicula keenani ),[113] и четыре блохи (Джеллисония sp., Polygenis roberti, Polygenis klagesi, и Polygenis dunni ) были найдены на T. talamancae в Панаме.[114] Г. аиткени также был обнаружен на этом виде в Колумбии.[105] Кроме того, сосущие вши Hoplopleura nesoryzomydis и Hoplopleura oryzomydis происходить на T. talamancae.[115]
Статус сохранения
Широко распространенный и распространенный вид, Transandinomys talamancae указан как "Наименьшие опасения " посредством Красный список МСОП. Встречается во многих охраняемые территории, хорошо переносит нарушенные среды обитания, и никаких серьезных угроз неизвестно.[2]
Примечания
- ^ Oryzomys megacephalus, как использовали Musser et al. (1998), была определена более широко, чем нынешняя Hylaeamys megacephalus (переведен в новый род Hylaeamys Векслера и его коллег в той же статье 2006 г., в которой Трансандиномис); по их определению, это произошло на всей территории Амазонии и на север до Тринидада и на юг до Парагвая.[37] Однако с тех пор популяции этого вида в западной Амазонии были разделены на другие виды, Hylaeamys perenensis. В следующих сравнениях, основанных на Musser et al. (1998), которые не разделяли их, а только H. megacephalus упоминается, но оба вида морфологически практически идентичны и, как известно, отличаются только размером, кариотип, и митохондриальная ДНК последовательности.[38]
- ^ Количество измеренных образцов.
- ^ 19 для длины хвоста.
- ^ Мужские добавочные железы T. talamancae, и ряда других сигмодонтинов, были описаны Voss и Linzey (1981). Их образец "Oryzomys capito"включены образцы из Панамы и Тринидада, представляющие Transandinomys talamancae и Hylaeamys megacephalusсоответственно, но их описание применимо к обоим.[74]
- ^ В одном образце не удалось определить, было ли FN 66 или 67; у остальных трех было 66.[78]
- ^ Сообщается как Oryzomys megacephalus по Musser et al. (1998), но только perenensis в настоящее время записывается из Колумбии.[87]
- ^ Этот вид был описан среди других по экземплярам "Оризомыс латицепс"из Рио-Кобарии, Департамент Араука[105] (фактически в Департамент Бояка );[106] Сан-Хуан Непомучено, Боливарское отделение;[105] Сокорре, верховья Рио-Сину, Боливар, Колумбия;[107] и Серро Асуль, Панама.[104] "Оризомыс латицепс"по определению 1960-х годов Transandinomys talamancae и никаких других видов, встречающихся в этом регионе;[108] кроме того, Musser et al. (1998) внесены в список Oryzomys talamancae для всех этих мест.[109]
Рекомендации
- ^ а б Гольдман, 1918 г., пластина IV
- ^ а б c Андерсон и др., 2017
- ^ Musser et al., 1998, рис. 66; Линарес, 1998, карта 134
- ^ Аллен, 1891, стр. 193
- ^ Аллен, 1899, стр. 208
- ^ Аллен, 1899, стр. 209
- ^ Аллен, 1899, стр. 210
- ^ Томас, 1900, стр. 272
- ^ Томас, 1901, стр. 252
- ^ Робинсон и Лион, 1901, стр. 142
- ^ Аллен, 1908, стр. 656
- ^ а б c d Weksler et al., 2006, стр. 25
- ^ Musser et al., 1998, стр. 273–274.
- ^ Аллен, 1891, стр. 193–194.
- ^ Аллен, 1899, стр. 208–210; Musser et al., 1998, стр. 273–274.
- ^ Томас, 1900, стр. 272–273.
- ^ Thomas, 1901, стр. 252–253.
- ^ Робинсон и Лион, 1901, стр. 142–143.
- ^ Аллен, 1908, стр. 656–657.
- ^ Гольдман, 1918, с. 71–74.
- ^ Гершковиц, 1960, стр. 544; Musser et al., 1998, стр. 179
- ^ Массер и Уильямс, 1985, стр. 9
- ^ Массер и Уильямс, 1985, стр. 7
- ^ Массер и Уильямс, 1985, стр. 13–14.
- ^ Musser et al., 1998, стр. 275–276.
- ^ Weksler, 2006, стр. 75–77.
- ^ Weksler, 2006, фиг. 34–39
- ^ Weksler et al., 2006, стр. 26
- ^ Векслер, 2006, с. 3
- ^ Мюссер и Карлтон, 2005
- ^ а б c d е ж грамм Гольдман, 1918, с. 73
- ^ а б Массер и Карлтон, 2005, стр. 1155
- ^ Массер и Уильямс, 1985, стр. 14; Musser et al., 1998, стр. 173
- ^ Musser et al., 1998, стр. 125
- ^ Musser et al., 1998, стр. 127
- ^ Musser et al., 1998, стр. 167, 169.
- ^ а б c Musser et al., 1998, стр. 169
- ^ Паттон и др., 2000, стр. 140
- ^ Массер и Уильямс, 1985, стр. 18; Musser et al., 1998, стр. 169
- ^ а б c d е ж Массер и Уильямс, 1985, стр. 14
- ^ Musser et al., 1998, стр. 127–128.
- ^ а б Musser et al., 1998, стр. 128–129.
- ^ а б c d е Musser et al., 1998, стр. 173
- ^ а б Флеминг, 1970, стр. 479
- ^ а б Гольдман, 1918, с. 71
- ^ Мюссер и Уильямс, 1985, таблица 2
- ^ а б c Musser et al., 1998, стр. 129
- ^ а б Musser et al., 1998, стр. 131
- ^ Гольдман, 1918, с. 71–72.
- ^ Гольдман, 1918, с. 72
- ^ Weksler et al., 2006, стр. 25; Musser et al., 1998, стр. 124
- ^ Линарес, 1998, стр. 287; Тирира, 2007, с. 200; Рид, 2009, стр. 208
- ^ Weksler, 2006, с. 17, 19.
- ^ Степпан, 1995 г., стол 5
- ^ Гольдман, 1918, с. 73; Массер и Уильямс, 1985, стр. 14
- ^ Weksler, 2006, стр. 31–32.
- ^ Векслер, 2006, с. 29, таблица 5
- ^ а б Musser et al., 1998, стр. 135
- ^ а б Musser et al., 1998, стр. 174
- ^ Массер и Уильямс, 1985, стр. 14; Weksler, 2006, стр. 34–35.
- ^ Гольдман, 1918, с. 73; Векслер, 2006, с. 35; Массер и Уильямс, 1985, стр. 14
- ^ Векслер, 2006, стр. 38–39.
- ^ а б Musser et al., 1998, стр. 140
- ^ Векслер, 2006, с. 41 год
- ^ Векслер, 2006, с. 42
- ^ Векслер, 2006, с. 43
- ^ Weksler, 2006, стр. 43–44.
- ^ Musser et al., 1998, стр. 140–141.
- ^ Weksler, 2006, стр. 42–43.
- ^ Гольдман, 1918 г., тарелка V
- ^ Векслер, 2006, с. 55
- ^ Weksler, 2006, стр. 55–56.
- ^ Weksler, 2006, стр. 56–57.
- ^ Векслер, 2006, с. 58, сноска 10
- ^ Weksler, 2006, стр. 57–58; Восс и Линзи, 1981, стр. 13
- ^ Гольдман, 1918 г., пластина VI
- ^ Musser et al., 1998, стр. 163
- ^ Musser et al., 1998, стр. 163–164.
- ^ Musser et al., 1998, стр. 164–165.
- ^ а б Musser et al., 1998, стр. 165
- ^ Musser et al., 1998, стр. 165–166.
- ^ Musser et al., 1998, стр. 125, 169, таблица 13
- ^ Musser et al., 1998, стр. 174; Паттон и др., 2000, стр. 140
- ^ а б Musser et al., 1998, стр. 158
- ^ а б c d Musser et al., 1998, стр. 157
- ^ Musser et al., 1998, стр. 157–158, сноска 9
- ^ Массер и Карлтон, 2005, стр. 1151, 1153
- ^ Рид, 2009, стр. 208; Тирира, 2007, с. 200
- ^ Флеминг, 1970; 1971 г.
- ^ Флеминг, 1971, стр. 5
- ^ Флеминг, 1971, стр. 60
- ^ Флеминг, 1971, стр. 50
- ^ Флеминг, 1971, стр. 22
- ^ Флеминг, 1971, стр. 23, рис. 6
- ^ а б c Линарес, 1998, стр. 288
- ^ Флеминг, 1971, стр. 40
- ^ Флеминг, 1971, стр. 65
- ^ Флеминг, 1971, таблица 11
- ^ Флеминг, 1971, стр. 41 год
- ^ Флеминг, 1971, стр. 43
- ^ Флеминг, 1971, стр. 29
- ^ Флеминг, 1971, стр. 32
- ^ Флеминг, 1971, стр. 48
- ^ а б Ли и Страндтманн, 1967, стр. 30
- ^ а б c Ли и Страндтманн, 1967, стр. 28
- ^ Musser et al., 1998, стр. 259
- ^ Ли и Страндтманн, 1967, стр. 29
- ^ Гершковиц, 1960, стр. 544; см. Musser et al. (1998) для обзора.
- ^ Musser et al., 1998, с. 150 (местонахождение 16), 151 (местонахождение 52, 53), 153 (местонахождение 68).
- ^ Типтон и др., 1966, стр. 42
- ^ Кларк, 1967
- ^ Бреннан и Рид, 1979, стр. 535
- ^ Бреннан и Юнкер, 1966, стр. 262
- ^ Типтон и Мендес, 1966, стр. 330
- ^ Дерден и Массер, 1994, стр. 30
Цитированная литература
- Аллен, Дж. 1891 г. Описания двух предполагаемых новых видов мышей из Коста-Рики и Мексики с замечаниями по Hesperomys melanophrys Coues. Труды Национального музея США 14 (850): 193–196.
- Аллен, Дж. 1899 г. Новые грызуны из Колумбии и Венесуэлы. Бюллетень Американского музея естественной истории 12: 195–218.
- Аллен, Дж. 1908 г. Млекопитающие из Никарагуа. Бюллетень Американского музея естественной истории 24: 647–670.
- Андерсон, Р.П .; Aguilera, M .; Gómez-Laverde, M .; Самудио, Р. Jr .; Пино, Дж. Л. (2017). "Transandinomys talamancae". Красный список видов, находящихся под угрозой исчезновения МСОП. 2017: e.T15615A22332803. Получено 2 января 2020.
- Бреннан, Дж. М., Рид, Дж. 1973 г. Неотропический род Айткениус: Три новых вида и другие записи из Венесуэлы (Acarina: Trombiculidae) (требуется подписка). Журнал паразитологии 59 (3): 531–535.
- Бреннан, Дж. М., Юнкер, К. Э. 1966. Чиггеры Панамы (Acarina: Trombiculidae). pp. 221–266 в Wenzel, R.L. and Tipton, V.J. (ред.). Эктопаразиты Панамы. Чикаго: Полевой музей естественной истории.
- Кларк, Г. 1967 г. Новые Speleognathinae из млекопитающих Центральной и Южной Америки (Acarina: Trombidiformes). Труды Вашингтонского гельминтологического общества 34: 240–243.
- Дерден, Л.А., Массер, Г.Г. 1994 г. Сосущие вши (Insecta, Anoplura) в мире: таксономический контрольный список с записями о млекопитающих-хозяевах и географическом распределении. Бюллетень Американского музея естественной истории 218: 1–90.
- Флеминг, Т. 1970 г. Заметки о фауне грызунов двух панамских лесов (требуется подписка). Журнал маммологии 51 (3): 473–490.
- Флеминг, Т. 1971 г. Популяционная экология трех видов неотропических грызунов. Разные публикации Зоологического музея Мичиганского университета 143: 1–77.
- Гольдман, Э.А. 1918 г. Рисовые крысы Северной Америки. Фауна Северной Америки 43: 1–100.
- Хэндли, C.O., младший, 1966. Контрольный список млекопитающих Панамы. стр. 753–795 в Wenzel, R.L. and Tipton, V.J. (ред.). Эктопаразиты Панамы. Чикаго: Полевой музей естественной истории, xii + 861 стр.
- Гершковиц, П. 1960 г. Млекопитающие северной Колумбии, предварительный отчет нет. 8. Древесные рисовые крысы, систематический пересмотр подрода. Oecomys, род Оризомыс. Труды Национального музея США 110: 513–568.
- Ли Д. и Страндтманн Р. В. 1967. Два новых вида Гигантолаэлапс (Acarina: Laelaptidae) с ключом к самкам (требуется подписка). Журнал Канзасского энтомологического общества 40 (1): 25–32.
- Линарес, О. 1998. Mamíferos de Venezuela. Каракас: Sociedad Conservacionista Audubon de Venezuela и British Petroleum, 691 стр. (На испанском языке). ISBN 980-6326-16-4
- Мюссер, Г.; Карлтон, доктор медицины (2005). «Надсемейство Muroidea». В Уилсон, Д.; Ридер, Д.М. (ред.). Виды млекопитающих мира: таксономический и географический справочник (3-е изд.). Издательство Университета Джона Хопкинса. п. 1155. ISBN 978-0-8018-8221-0. OCLC 62265494.
- Мюссер, Г. и Уильямс, М. 1985 г. Систематические исследования грызунов оризомиинов (Muridae): определения Oryzomys villosus и Oryzomys talamancae. Американский музей Novitates 2810: 1–22.
- Массер, Г.Г., Карлтон, доктор медицины, братья Э.М. и Гарднер, А.Л. 1998. Систематические исследования грызунов оризомиинов (Muridae: Sigmodontinae): диагностика и распространение видов, ранее отнесенных к Оризомыс "капито". Бюллетень Американского музея естественной истории 236: 1–376.
- Паттон, Дж. Л., да Силва, М. Н. Ф. и Малькольм, Дж. Р. 2000. Млекопитающие Рио-Джуруа и эволюционное и экологическое разнообразие Амазонии. Бюллетень Американского музея естественной истории 244: 1–306.
- Рид, Ф. 2009. Полевой справочник по млекопитающим Центральной Америки и Юго-Восточной Мексики. 2-е издание. Oxford University Press, США, 346 стр. ISBN 978-0-19-534322-9
- Робинсон, В. и Лайон, М.В., мл. 1901. Аннотированный список млекопитающих, собранных в окрестностях Ла Гуайра, Венесуэла.. Труды Национального музея США 24 (1246): 135–162.
- Томас, О. 1900. Описание новых неотропических млекопитающих. Анналы и журнал естественной истории (7) 5: 269–274.
- Томас, О. 1901. Новые неотропические млекопитающие с примечанием о видах Reithrodon. Летопись и журнал естественной истории (7) 8: 246–255.
- Типтон, В.Дж. and Méndez, E. 1966. Блохи (Siphonaptera) Панамы. стр. 289–385 в Wenzel, R.L. and Tipton, V.J. (ред.). Эктопаразиты Панамы. Чикаго: Полевой музей естественной истории.
- Типтон, В.Дж., Альтман, Р.М. и Кинан, К. 1966 г. Клещи подсемейства Laelaptinae в Панаме (Acarina: Laelaptidae). стр. 23–82 в Wenzel, R.L. and Tipton, V.J. (ред.). Эктопаразиты Панамы. Чикаго: Полевой музей естественной истории.
- Тирира, Д. 2007. Guia de campo de los mamíferos del Ecuador. Кито: Ediciones Murciélago Blanco, publicación especial sobre los mamíferos del Ecuador 6, 576 стр. (На испанском языке). ISBN 9978-44-651-6
- Векслер М. 2006. Филогенетические взаимоотношения грызунов оризомиинов (Muroidea: Sigmodontinae): отдельный и комбинированный анализ морфологических и молекулярных данных. Бюллетень Американского музея естественной истории 296: 1–149.
- Векслер М., Персекильо А. и Восс, Р. 2006 г. Десять новых родов грызунов оризомиинов (Cricetidae: Sigmodontinae). Американский музей Novitates 3537: 1–29.
внешняя ссылка
 СМИ, связанные с Oryzomys talamancae в Wikimedia Commons
СМИ, связанные с Oryzomys talamancae в Wikimedia Commons Данные, относящиеся к Transandinomys talamancae в Wikispecies
Данные, относящиеся к Transandinomys talamancae в Wikispecies