Polyomaviridae - Polyomaviridae
| Polyomaviridae | |
|---|---|
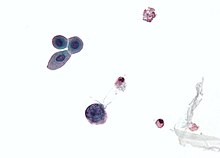 | |
| Микрофотография показывая полиомавирус инфицированная ячейка - большая (синяя) ячейка внизу в центре слева. Цитологический образец мочи. | |
| Классификация вирусов | |
| (без рейтинга): | Вирус |
| Область: | Моноднавирия |
| Королевство: | Shotokuvirae |
| Тип: | Cossaviricota |
| Учебный класс: | Papovaviricetes |
| Заказ: | Сеполивиралес |
| Семья: | Polyomaviridae |
| Роды | |
Polyomaviridae это семья вирусы чей естественный хозяева в первую очередь млекопитающие и птицы.[1][2] По состоянию на 2019 год насчитывается четыре признанных рода и 102 вида, девять из которых не относятся к роду.[3] Известно, что 14 видов заражают людей, в то время как другие, такие как Обезьяний вирус 40, были идентифицированы у людей в меньшей степени.[4][5] Большинство этих вирусов очень распространены и обычно бессимптомны в большинстве изученных популяций людей.[6][7] BK вирус связан с нефропатия в почечный трансплантат пациенты с трансплантацией непочечных твердых органов,[8][9] JC вирус с прогрессирующая мультифокальная лейкоэнцефалопатия,[10] и Вирус клетки Меркель с Рак из клеток Меркеля.[11]
Структура и геном

Полиомавирусы без оболочки двухцепочечная ДНК вирусы с циркулярным геномы около 5000 пар оснований. Геном упакован в вирусный капсид около 40-50 нанометры в диаметре, который икосаэдр по форме (симметрия Т = 7).[2][12] Капсид состоит из 72 пентамеров. капсомеры белка, называемого VP1, способный к самосборке в замкнутый икосаэдр;[13] каждый пентамер VP1 связан с одной молекулой одного из двух других капсидных белков, VP2 или VP3.[5]

Геном типичного полиомавируса кодирует от 5 до 9 белки, разделенный на два транскрипционный регионы называются ранними и поздними регионами из-за времени во время заражения, в которое они транскрибируются. Каждая область транскрибируется клеткой-хозяином РНК-полимераза II как сингл пре-мессенджер РНК содержащий несколько генов. Ранняя область обычно кодирует два белка, малый и большой опухолевые антигены, продуцируемые альтернативное сращивание. Поздняя область содержит три структурных белка капсида VP1, VP2 и VP3, продуцируемые альтернативным способом. переводной стартовые сайты. Дополнительные гены и другие вариации на эту тему присутствуют в некоторых вирусах: например, полиомавирусы грызунов имеют третий белок, называемый средний опухолевый антиген в ранней области, что чрезвычайно эффективно для индукции клеточная трансформация; SV40 имеет дополнительный капсидный белок VP4; в некоторых примерах есть дополнительный регуляторный белок, называемый агнопротеин выражено из позднего региона. В геноме также есть некодирование контрольный или регуляторный регион, содержащий ранние и поздние регионы ' промоутеры, стартовые сайты транскрипции и начало репликации.[2][12][5][15]
| Род | Структура | Симметрия | Капсид | Геномное расположение | Геномная сегментация |
|---|---|---|---|---|---|
| Полиомавирус | Икосаэдр | Т = 7 | Без оболочки | Круговой | Одночастный |
Репликация и жизненный цикл
Жизненный цикл полиомавируса начинается с попадания в клетка-хозяин. Клеточные рецепторы полиомавирусов: сиаловая кислота остатки гликаны, обычно ганглиозиды. Присоединение полиомавирусов к клеткам-хозяевам опосредуется связыванием VP1 к сиалированным гликанам на поверхности клетки.[2][12][15][16] В некоторых конкретных вирусах происходят дополнительные взаимодействия клеточной поверхности; например, JC вирус считается, что требуется взаимодействие с Рецептор 5HT2A и Вирус клетки Меркель с гепарансульфат.[15][17] Однако в целом взаимодействия вируса с клеткой опосредуются часто встречающимися молекулами на поверхности клетки и поэтому, вероятно, не являются основным фактором, влияющим на наблюдаемый тип клеток отдельных вирусов. тропизм.[15] После связывания с молекулами на поверхности клетки вирион эндоцитозированный и входит в эндоплазматический ретикулум - поведение, уникальное среди известных вирусов без оболочки[18] - где структура вирусного капсида может быть нарушена действием клетки-хозяина дисульфид изомераза ферменты.[2][12][19]
Детали перехода к ядру неясны и могут различаться для разных полиомавирусов. Часто сообщалось, что интактная, хотя и искаженная, частица вириона высвобождается из эндоплазматического ретикулума в цитоплазму клетки, где геном высвобождается из капсида, возможно, из-за низкого кальций концентрация в цитоплазме.[18] Как экспрессия вирусных генов, так и репликация вирусного генома происходят в ядро с использованием механизмов клетки-хозяина. Ранние гены, включающие как минимум небольшой опухолевый антиген (Стоять большой опухолевый антиген (LT) - выражаются первыми, из единственного альтернативно сращенный информационная РНК прядь. Эти белки служат для манипулирования хозяином клеточный цикл - нарушение регуляции перехода от Фаза G1 к Фаза S, когда геном клетки-хозяина реплицируется - потому что для репликации вирусного генома необходим механизм репликации ДНК клетки-хозяина.[2][12][15] Точный механизм этого нарушения регуляции зависит от вируса; Например, SV40 LT может напрямую связываться с клеткой-хозяином p53, но мышиный полиомавирус LT нет.[20] LT индуцирует репликацию ДНК из некодирующей контрольной области вирусного генома (NCCR), после чего экспрессия ранней мРНК снижается и начинается экспрессия поздней мРНК, которая кодирует белки вирусного капсида.[19] Когда эти взаимодействия начинаются, LT, принадлежащие нескольким полиомавирусам, в том числе Полиомавирус клеток Меркеля, присутствует онкогенный потенциал.[21]Было описано несколько механизмов для регуляции перехода от ранней к поздней экспрессии гена, включая участие белка LT в репрессии раннего промотора,[19] экспрессия незавершенных поздних мРНК с расширениями, комплементарными ранней мРНК,[15] и проявление регуляторных микроРНК.[15]Экспрессия поздних генов приводит к накоплению белков вирусного капсида в цитоплазме клетки-хозяина. Компоненты капсида проникают в ядро для инкапсидации новой вирусной геномной ДНК. Новые вирионы могут быть собраны в вирусные фабрики.[2][12] Механизм высвобождения вируса из клетки-хозяина варьируется среди полиомавирусов; некоторые экспрессируют белки, которые облегчают выход из клетки, такие как агнопротеин или же VP4.[19] В некоторых случаях высокие уровни инкапсидированного вируса приводят к тому, что клетки лизис, высвобождая вирионы.[15]
| Род | Детали хоста | Тканевый тропизм | Детали входа | Детали выпуска | Сайт репликации | Сайт сборки | Передача инфекции |
|---|---|---|---|---|---|---|---|
| Полиомавирус | Млекопитающие; птицы | Дыхательная система; почки, мозг | Эндоцитоз клеточных рецепторов | Лизис | Ядро | Ядро | Орально-фекальный |
Вирусные белки
Опухолевые антигены
В большой опухолевый антиген играет ключевую роль в регулировании жизненного цикла вируса, связываясь с вирусным источником репликации ДНК, где он способствует синтезу ДНК. Также, поскольку полиомавирус полагается на аппарат клетки-хозяина для репликации, клетка-хозяин должна находиться в s-фазе, чтобы это началось. Благодаря этому большой Т-антиген также модулирует клеточные сигнальные пути, чтобы стимулировать развитие клеточного цикла, связываясь с рядом клеточных контрольных белков.[22] Это достигается за счет двухсторонней атаки: подавления опухолевых генов p53 и членов ретинобластома (pRB) семья,[23] и стимуляция путей роста клеток путем связывания клеточной ДНК, АТФаза-геликаза, ассоциации ДНК-полимеразы-α и связывания факторов комплекса преинициации транскрипции.[24] Эта ненормальная стимуляция клеточного цикла - мощная сила для онкогенной трансформации.
В небольшой опухолевый антиген белок также способен активировать несколько клеточных путей, которые стимулируют пролиферацию клеток. Малые Т-антигены полиомавирусов обычно нацелены на протеинфосфатазу 2А (PP2A ),[25] ключевой мультисубъединичный регулятор множества путей, включая Акт путь митоген-активируемой протеинкиназы (MAPK) и путь стресс-активируемой протеинкиназы (SAPK).[26][27] Полиомавирус клеток Меркеля малый Т-антиген кодирует уникальный домен, называемый доменом стабилизации LT (LSD), который связывается и ингибирует FBXW7 E3 лигаза регулирующие как клеточные, так и вирусные онкопротеины.[28] В отличие от SV40, малый Т-антиген MCV непосредственно трансформирует клетки грызунов in vitro.[29]
В средний опухолевый антиген используется в модельные организмы разработан для изучения рака, например ММТВ-ПыМТ система, в которой средний Т соединен с ММТВ промоутер. Там он функционирует как онкоген, а ткань, в которой развивается опухоль, определяется промотором MMTV.
Капсидные белки
Капсид полиомавируса состоит из одного основного компонента, главный капсидный белок VP1, и один или два второстепенных компонента, минорные капсидные белки VP2 и VP3. VP1 пентамеры сформировать закрытый икосаэдр вирусный капсид, а внутри капсида каждый пентамер связан с одной молекулой либо VP2, либо VP3.[5][30] Некоторые полиомавирусы, такие как Полиомавирус клеток Меркеля, не кодируют и не выражают VP3.[31] Белки капсида экспрессируются из поздней области генома.[5]
Агнопротеин
В агнопротеин представляет собой небольшой многофункциональный фосфопротеин, обнаруженный в поздней кодирующей части генома некоторых полиомавирусов, в первую очередь BK вирус, JC вирус, и SV40. Он важен для пролиферации вирусов, которые его экспрессируют, и считается, что он участвует в регуляции жизненного цикла вируса, в частности, репликации и выхода вируса из клетки-хозяина, но точные механизмы неясны.[32][33]
Таксономия
Полиомавирусы относятся к группе I (вирусы дцДНК). Классификация полиомавирусов была предметом нескольких предложенных изменений по мере обнаружения новых членов группы. Ранее полиомавирусы и папилломавирусы, которые имеют много общих структурных особенностей, но имеют очень разные геномные организации, были классифицированы вместе в уже устаревшее семейство Papovaviridae.[34] (Название Papovaviridae происходит от трех сокращений: Па для Папилломавирус, Po для Полиомавирус, и Va для «вакуолизации».)[35] Полиомавирусы были разделены на три основных клады (то есть генетически родственные группы): кладу SV40, кладу птиц и кладу мышиных полиомавирусов.[36] Последующая предложенная реклассификация Международный комитет по таксономии вирусов (ICTV) рекомендовала разделить семейство Polyomaviridae на три рода:[37]
- Род Ортополиомавирус (типовой вид SV40 )
- Род Вукиполёмовирус (типовой вид Полиомавирус KI )
- Род Avipolyomavirus (типовой вид Птичий полиомавирус )
Текущая система классификации ICTV распознает четыре рода и 102 вида, из которых девять не могут быть отнесены к роду. Эта система сохраняет различие между птичьим вирусом и вирусом млекопитающих, группируя птичью субпопуляцию в род Гаммаполиомавирус.[3]
- Род Альфаполиомавирус, типовой вид Полиомавирус Mus musculus 1 (мышиный полиомавирус)
- Род Бетаполиомавирус, типовой вид Полиомавирус Macaca mulatta 1 (SV40)
- Род Дельтаполиомавирус, типовой вид Полиомавирус человека 6
- Род Гаммаполиомавирус, типовой вид Полиомавирус Aves 1
Следующие виды не отнесены к роду:[3]
- Полиомавирус Ailuropoda melanoleuca 1
- Полиомавирус Bos taurus 1
- Полиомавирус Centropristis striata 1
- Полиомавирус Delphinus delphis 1
- Процион лоторный полиомавирус 2
- Полиомавирус 1 Rhynchobatus djiddensis
- Полиомавирус Sparus aurata 1
- Полиомавирус Trematomus bernacchii 1
- Полиомавирус Trematomus pennellii 1
Описание дополнительных вирусов продолжается. К ним относятся полиомавирус калана 1[38] и полиомавирус альпаки[39] Другой вирус - полиомавирус гигантской панды 1.[40] Другой вирус был описан от грызунов сигмодонтинов.[41] Другой - полиомавирус 1 бурозубки - был описан у землероек.[42]
Полиомавирусы человека
Большинство полиомавирусов не инфицируют людей. Из полиомавирусов, каталогизированных по состоянию на 2017 год, в общей сложности 14 были известны с человеческими хозяевами.[4] Однако некоторые полиомавирусы связаны с болезнями человека, особенно в с ослабленным иммунитетом лиц. MCV сильно отличается от других полиомавирусов человека и наиболее близок к полиомавирусу мышей. Полиомавирус, связанный с Trichodysplasia spinulosa (TSV) отдаленно связан с MCV. Два вируса - HPyV6 и HPyV7 - наиболее тесно связаны с вирусами KI и WU, в то время как HPyV9 наиболее тесно связан с лимфотропным полиомавирусом африканских зеленых обезьян (LPV).
Описан четырнадцатый вирус.[43] Полиомавирус Lyon IARC относится к полиомавирусу енота.
Список полиомавирусов человека
Следующие 14 полиомавирусов с человеческими хозяевами были идентифицированы, и их геномы последовательность по состоянию на 2017 год:[4]
| Разновидность | Предлагаемый род | Название вируса | Сокращение | NCBI RefSeq | Год открытия | Клинический коррелят (если есть) | Рекомендации |
|---|---|---|---|---|---|---|---|
| Полиомавирус человека 5 | Альфа | Полиомавирус клеток Меркеля | MCPyV | NC_010277 | 2008 | Рак из клеток Меркеля[5] | [44][11][45] |
| Полиомавирус человека 8 | Альфа | Полиомавирус Trichodysplasia spinulosa | TSPyV | NC_014361 | 2010 | Спинулезная триходисплазия[5] | [46][47] |
| Полиомавирус человека 9 | Альфа | Полиомавирус человека 9 | HPyV9 | NC_015150 | 2011 | Никто не известен | [48] |
| Полиомавирус человека 12 | Альфа | Полиомавирус человека 12 | HPyV12 | NC_020890 | 2013 | Никто не известен | [49] |
| Полиомавирус человека 13 | Альфа | Полиомавирус Нью-Джерси | NJPyV | NC_024118 | 2014 | Никто не известен | [50] |
| Полиомавирус человека 1 | Бета | BK полиомавирус | BKPyV | NC_001538 | 1971 | Связанный с полиомавирусом нефропатия; геморрагический цистит[5] | [51] |
| Полиомавирус человека 2 | Бета | Полиомавирус JC | JCPyV | NC_001699 | 1971 | Прогрессирующая мультифокальная лейкоэнцефалопатия[5] | [52] |
| Полиомавирус человека 3 | Бета | Полиомавирус KI | KIPyV | NC_009238 | 2007 | Никто не известен | [53] |
| Полиомавирус человека 4 | Бета | Полиомавирус WU | WUPyV | NC_009539 | 2007 | Никто не известен | [14] |
| Полиомавирус человека 6 | Дельта | Полиомавирус человека 6 | HPyV6 | NC_014406 | 2010 | Зуд и дискератотический дерматоз, ассоциированный с HPyV6 (H6PD)[54] | [31] |
| Полиомавирус человека 7 | Дельта | Полиомавирус человека 7 | HPyV7 | NC_014407 | 2010 | Эпителиальная гиперплазия, связанная с HPyV7[54][55][56] | [31] |
| Полиомавирус человека 10 | Дельта | Полиомавирус MW | MWPyV | NC_018102 | 2012 | Никто не известен | [57][58][59] |
| Полиомавирус человека 11 | Дельта | Полиомавирус STL | STLPyV | NC_020106 | 2013 | Никто не известен | [60] |
| Полиомавирус человека 14 | Альфа | Полиомавирус Lyon IARC | LIPyV | NC_034253.1 | 2017 | Никто не известен | [61][62] |
Дельтаполиомавирус содержит только четыре вируса человека, представленных в таблице выше, с типовым видом полиомавируса человека 6. Группы альфа и бета содержат вирусы, поражающие различных млекопитающих. Группа «Гамма» содержит вирусы птиц.[4] Клинически значимые ассоциации болезни показаны только там, где ожидается причинность.[5][63]
Антитела к лимфотропному полиомавирусу обезьян были обнаружены у людей, что позволяет предположить, что этот вирус - или близкородственный вирус - может инфицировать людей.[64]
Клиническая значимость
Все полиомавирусы очень распространены среди детей и молодых взрослых.[65] Большинство этих инфекций практически не вызывают симптомов. Эти вирусы, вероятно, сохраняются на протяжении всей жизни почти у всех взрослых. Заболевания, вызванные инфекциями полиомавируса человека, наиболее распространены среди с ослабленным иммунитетом люди; ассоциации болезней включают BK вирус с нефропатия в почечный трансплантат пациенты с трансплантацией солидных органов, не относящиеся к почкам,[8][9] JC вирус с прогрессирующая мультифокальная лейкоэнцефалопатия,[10] и Вирус клетки Меркель (MCV) с Рак из клеток Меркеля.[11]
SV40
SV40 реплицируется в почках обезьяны не вызывая заболевания, но может вызвать рак у грызунов в лабораторных условиях. В 1950-х и начале 1960-х годов более 100 миллионов человек могли подвергнуться воздействию SV40 из-за ранее не обнаруженного заражения SV40. вакцина от полиомиелита, что вызывает опасения по поводу возможности того, что вирус может вызвать заболевание у людей.[66][67] Хотя сообщалось, что он присутствует при некоторых раковых заболеваниях человека, включая опухоли головного мозга, костные опухоли, мезотелиомы, и неходжкинские лимфомы,[68] точное обнаружение часто затрудняется высоким уровнем перекрестной реактивности SV40 с широко распространенными полиомавирусами человека.[67] Большинство вирусологов отвергают SV40 как причину рака человека.[66][69][70]
Диагностика
Диагноз полиомавируса почти всегда ставится после первичного инфицирования, поскольку он протекает бессимптомно или субклинически. Анализы антител обычно используются для определения наличия антител против отдельных вирусов.[71] Чтобы отличить очень похожие полиомавирусы, часто необходимы конкурентные анализы.[72]
В случаях прогрессирующей мультифокальной лейкоэнцефалопатии (ПМЛ) перекрестно-реактивные антитела к Т-антигену SV40 (обычно Pab419) используются для окрашивания тканей непосредственно на наличие Т-антигена вируса JC. ПЦР может использоваться при биопсии ткани или спинномозговая жидкость для амплификации ДНК полиомавируса. Это позволяет не только обнаруживать полиомавирус, но и определять его подтип.[73]
Для диагностики реактивации полиомавируса при полиомавирусной нефропатии (ПВН) используются три основных диагностических метода: цитология мочи, количественное определение вирусной нагрузки как в моче, так и в крови и почечная биопсия.[71]Реактивация полиомавируса в почках и мочевыводящих путях вызывает выделение инфицированных клеток, вирионов и / или вирусных белков с мочой. Это позволяет цитологическому анализу мочи исследовать эти клетки, что, если есть включение полиомавируса в ядро, является диагностическим признаком инфекции.[74] Кроме того, поскольку моча инфицированного человека будет содержать вирионы и / или вирусную ДНК, количественное определение вирусной нагрузки может быть выполнено с помощью ПЦР.[75] То же самое и с кровью.
Биопсия почек также может быть использована, если два только что описанных метода не позволяют сделать окончательный результат или если требуется определенная вирусная нагрузка на почечную ткань. Подобно цитологии мочи, почечные клетки исследуются под световой микроскопией на наличие полиомавируса в ядре, а также на лизис клеток и вирусные частицы во внеклеточной жидкости. Вирусная нагрузка, как и прежде, также измеряется с помощью ПЦР.[нужна цитата ]
Окрашивание тканей с использованием моноклональных антител против Т-антигена MCV показывает полезность для дифференциации карциномы из клеток Меркеля от других небольших круглоклеточных опухолей.[76] Были разработаны анализы крови для выявления антител к MCV, которые показывают, что инфекция вирусом широко распространена, хотя пациенты с карциномой из клеток Меркеля имеют исключительно более высокие ответы антител, чем лица, инфицированные бессимптомно.[7][77][78][79]
Использование для отслеживания миграции людей
Вирус JC является многообещающим генетическим маркером эволюции и миграции человека.[80] Он переносится 70–90 процентами людей и обычно передается от родителей к потомству. Этот метод не кажется надежным для отслеживания недавнее африканское происхождение современного человека.
История
Полиомавирус мыши был первым обнаруженным полиомавирусом, о котором сообщил Людвик Гросс в 1953 г. как вытяжка из мыши лейкемии способный вызвать околоушной железы опухоли.[81] Возбудитель был идентифицирован как вирус Сара Стюарт и Бернис Эдди, в честь которого когда-то называлась «SE полиома».[82][83][84] Термин «полиома» относится к способности вирусов продуцировать множественные (поли) опухоли (-омы) при определенных условиях. Название критиковали как «языковой бутерброд без мяса» («без мяса», потому что оба морфемы в «полиоме» - аффиксы) дающие мало информации о биологии вирусов; Фактически, последующие исследования показали, что большинство полиомавирусов редко вызывают клинически значимое заболевание у их организмов-хозяев в естественных условиях.[85]
По состоянию на 2017 год были идентифицированы и секвенированы десятки полиомавирусов, заражающих в основном птиц и млекопитающих. Известно, что два полиомавируса поражают рыб: черный морской окунь[86] и морской лещ.[87] Известно, что всего четырнадцать полиомавирусов инфицируют людей.[4]
Рекомендации
- ^ Moens U, Calvignac-Spencer S, Lauber C, Ramqvist T, Feltkamp MC, Daugherty MD, Verschoor EJ, Ehlers B (июнь 2017 г.). "Профиль таксономии вирусов ICTV: Polyomaviridae". Журнал общей вирусологии. 98 (6): 1159–1160. Дои:10.1099 / jgv.0.000839. ЧВК 5656788. PMID 28640744.
- ^ а б c d е ж грамм "Отчет ICTV о Polyomaviridae".
- ^ а б c «Таксономия вирусов: выпуск 2019 г.». talk.ictvonline.org. Международный комитет по таксономии вирусов. Получено 26 апреля 2020.
- ^ а б c d е Calvignac-Spencer S, Feltkamp MC, Daugherty MD, Moens U, Ramqvist T, Johne R, Ehlers B (июнь 2016 г.). «Обновление таксономии семейства Polyomaviridae». Архив вирусологии. 161 (6): 1739–50. Дои:10.1007 / s00705-016-2794-у. PMID 26923930.
- ^ а б c d е ж грамм час я j ДеКаприо Дж. А., Гарси Р. Л. (апрель 2013 г.). «Рог изобилия полиомавирусов человека». Обзоры природы. Микробиология. 11 (4): 264–76. Дои:10.1038 / nrmicro2992. ЧВК 3928796. PMID 23474680.
- ^ Госсай А., Ватербур Т., Нельсон Х. Х., Мишель А., Уиллхаук-Флекенштейн М., Фарзан С.Ф., Хоен А.Г., Кристенсен Б.К., Келси К.Т., Марсит С.Дж., Павлита М., Карагас М.Р. «Сероэпидемиология полиомавирусов человека в популяции США». Американский журнал эпидемиологии. 183 (1): 61–9. Дои:10.1093 / aje / kwv155. ЧВК 5006224. PMID 26667254.
- ^ а б Кин Дж. М., Рао С., Ван М., Гарси Р. Л. (март 2009 г.). «Сероэпидемиология полиомавирусов человека». Патогены PLOS. 5 (3): e1000363. Дои:10.1371 / journal.ppat.1000363. ЧВК 2655709. PMID 19325891.
- ^ а б Jamboti JS (август 2016 г.). «BK-вирусная нефропатия у реципиентов почечного трансплантата». Нефрология. 21 (8): 647–54. Дои:10.1111 / неп.12728. PMID 26780694.
- ^ а б Куппачи С., Каур Д., Холанда Д. Г., Томас С. П. (апрель 2016 г.). «Инфекция вирусом полиомы ВК и заболевание почек при трансплантации непочечных твердых органов». Клинический журнал почек. 9 (2): 310–8. Дои:10.1093 / ckj / sfv143. ЧВК 4792618. PMID 26985385.
- ^ а б Аданг Л., Бергер Дж. (2015). «Прогрессирующая мультифокальная лейкоэнцефалопатия». F1000 Исследования. 4: 1424. Дои:10.12688 / f1000research.7071.1. ЧВК 4754031. PMID 26918152.
- ^ а б c Фэн Х, Шуда М., Чанг Й, Мур П.С. (февраль 2008 г.). «Клональная интеграция полиомавируса в карциному из клеток Меркеля человека». Наука. 319 (5866): 1096–100. Bibcode:2008Sci ... 319.1096F. Дои:10.1126 / science.1152586. ЧВК 2740911. PMID 18202256.
- ^ Салунке Д.М., Каспар Д.Л., Гарси Р.Л. (сентябрь 1986 г.). «Самосборка очищенного капсидного белка полиомавируса VP1». Клетка. 46 (6): 895–904. Дои:10.1016/0092-8674(86)90071-1. PMID 3019556.
- ^ а б Гейнор А.М., Ниссен, доктор медицины, Вайли Д.М., Маккей И.М., Ламберт С.Б., Ву Г., Бреннан, округ Колумбия, Сторч Г.А., Слоутс Т.П., Ван Д. (май 2007 г.). «Выявление нового полиомавируса у пациентов с острыми респираторными инфекциями». Патогены PLOS. 3 (5): e64. Дои:10.1371 / journal.ppat.0030064. ЧВК 1864993. PMID 17480120.
- ^ а б c d е ж грамм час Международное агентство по изучению рака (2013 г.). «Введение в полиомавирусы» (PDF). Монографии МАИР по оценке канцерогенных рисков для человека. 104: 121–131.
- ^ а б Buch MH, Liaci AM, O'Hara SD, Garcea RL, Neu U, Stehle T (октябрь 2015 г.). «Структурный и функциональный анализ капсидных белков мышиного полиомавируса устанавливает детерминанты распознавания лиганда и патогенности». Патогены PLOS. 11 (10): e1005104. Дои:10.1371 / journal.ppat.1005104. ЧВК 4608799. PMID 26474293.
- ^ Шовальтер Р.М., Пастрана Д.В., Бак CB (июль 2011 г.). «Гликозаминогликаны и сиалированные гликаны последовательно способствуют проникновению инфекции полиомавируса клеток Меркеля». Патогены PLOS. 7 (7): e1002161. Дои:10.1371 / journal.ppat.1002161. ЧВК 3145800. PMID 21829355.
- ^ а б Иноуэ Т., Цай Б. (январь 2013 г.). «Как вирусы используют эндоплазматический ретикулум для проникновения, репликации и сборки». Перспективы Колд-Спринг-Харбор в биологии. 5 (1): a013250. Дои:10.1101 / cshperspect.a013250. ЧВК 3579393. PMID 23284050.
- ^ а б c d Gjoerup O, Chang Y (2010). «Обновленная информация о полиомавирусах и раке человека». Достижения в исследованиях рака. 106: 1–51. Дои:10.1016 / S0065-230X (10) 06001-X. ISBN 9780123747716. PMID 20399955.
- ^ Андраби С., Хван Дж. Х., Чхве Дж. К., Робертс Т.М., Шаффхаузен Б.С. (2011). «Сравнение полиомавируса мыши и обезьяньего вируса 40 показывает существенные различия в функции малого Т-антигена». Журнал вирусологии. 85 (20): 10649–10658. Дои:10.1128 / JVI.05034-11. ЧВК 3187521. PMID 21835797.
- ^ Rotondo JC, Bononi I, Puozzo A, Govoni M, Foschi V, Lanza G, Gafà R, Gaboriaud P, Touzé FA, Selvatici R, Martini F, Tognon M (июль 2017 г.). «Клеточные карциномы Меркеля, возникающие у пациентов с аутоиммунным заболеванием, леченных биологическими препаратами, включая анти-TNF». Клинические исследования рака. 23 (14): 3929–3934. Дои:10.1158 / 1078-0432.CCR-16-2899. PMID 28174236.
- ^ Белый МК, Гордон Дж., Рейсс К., Дель Валле Л., Кроул С., Джордано А., Дарбинян А., Халили К. (декабрь 2005 г.). «Полиомавирусы человека и опухоли головного мозга». Исследование мозга. Обзоры исследований мозга. 50 (1): 69–85. Дои:10.1016 / j.brainresrev.2005.04.007. PMID 15982744.
- ^ Kazem S, van der Meijden E, Wang RC, Rosenberg AS, Pope E, Benoit T., Fleckman P, Feltkamp MC (2014). «Trichodysplasia spinulosa, связанная с полиомавирусом, включает гиперпролиферацию, фосфорилирование pRB и активацию p16 и p21». PLOS One. 9 (10): e108947. Bibcode:2014PLoSO ... 9j8947K. Дои:10.1371 / journal.pone.0108947. ЧВК 4188587. PMID 25291363.
- ^ Келли В.Л., Георгопулос С. (апрель 1997 г.). «Общий экзон T / t обезьяньего вируса 40, полиомавируса JC и T-антигенов BK может функционально заменять J-домен молекулярного шаперона DnaJ Escherichia coli». Труды Национальной академии наук Соединенных Штатов Америки. 94 (8): 3679–84. Bibcode:1997PNAS ... 94.3679K. Дои:10.1073 / пнас.94.8.3679. ЧВК 20500. PMID 9108037.
- ^ Паллас, округ Колумбия, Шахрик Л.К., Мартин Б.Л., Ясперс С., Миллер Т.Б., Браутиган Д.Л., Робертс TM (январь 1990 г.). «Малый и средний Т-антигены полиомы и малый Т-антиген SV40 образуют стабильные комплексы с протеинфосфатазой 2А». Клетка. 60 (1): 167–76. Дои:10.1016 / 0092-8674 (90) 90726-у. PMID 2153055.
- ^ Зонтаг Э., Федоров С., Камибаяши С., Роббинс Д., Кобб М., Мамби М. (декабрь 1993 г.). «Взаимодействие небольшого опухолевого антигена SV40 с протеинфосфатазой 2A стимулирует путь киназы карты и индуцирует пролиферацию клеток». Клетка. 75 (5): 887–97. Дои:10.1016 / 0092-8674 (93) 90533-В. PMID 8252625.
- ^ Ватанабе Г., Хоу А., Ли Р.Дж., Альбанезе С., Шу И.В., Карнезис А.Н., Зон Л., Кириакис Дж., Рунделл К., Пестелл Р.Г. (ноябрь 1996 г.). «Индукция циклина D1 малым опухолевым антигеном вируса обезьяны 40». Труды Национальной академии наук Соединенных Штатов Америки. 93 (23): 12861–6. Bibcode:1996PNAS ... 9312861W. Дои:10.1073 / пнас.93.23.12861. ЧВК 24011. PMID 8917510.
- ^ Квун Х.Дж., Шуда М., Фенг Х., Камачо С.Дж., Мур П.С., Чанг Й. (август 2013 г.). «Малый Т-антиген полиомавируса клеток Меркеля контролирует репликацию вируса и экспрессию онкобелка, воздействуя на клеточную убиквитинлигазу SCFFbw7». Клеточный хозяин и микроб. 14 (2): 125–35. Дои:10.1016 / j.chom.2013.06.008. ЧВК 3764649. PMID 23954152.
- ^ Шуда М., Квун Х.Дж., Фенг Х., Чанг Й., Мур П.С. (сентябрь 2011 г.). «Малый Т-антиген полиомавируса клеток Меркеля человека представляет собой онкопротеин, нацеленный на регулятор трансляции 4E-BP1». Журнал клинических исследований. 121 (9): 3623–34. Дои:10.1172 / JCI46323. ЧВК 3163959. PMID 21841310.
- ^ Чен XS, Стеле Т., Харрисон СК (июнь 1998 г.). «Взаимодействие внутреннего белка полиомавируса VP2 с основным капсидным белком VP1 и последствия для участия VP2 в проникновении вируса». Журнал EMBO. 17 (12): 3233–40. Дои:10.1093 / emboj / 17.12.3233. ЧВК 1170661. PMID 9628860.
- ^ а б c Шовальтер Р.М., Пастрана Д.В., Памфри К.А., Мойер А.Л., Бак CB (июнь 2010 г.). «Полиомавирус клеток Меркеля и два ранее неизвестных полиомавируса хронически выделяются с кожи человека». Клеточный хозяин и микроб. 7 (6): 509–15. Дои:10.1016 / j.chom.2010.05.006. ЧВК 2919322. PMID 20542254.
- ^ Сарыер И.К., Сарибас А.С., Белый МК, Сафак М (май 2011 г.). «Инфекция агнопротеин-отрицательными мутантами полиомавируса JC и SV40 приводит к высвобождению вирионов, у которых в основном отсутствует ДНК». Журнал вирусологии. 8: 255. Дои:10.1186 / 1743-422X-8-255. ЧВК 3127838. PMID 21609431.
- ^ Сарибас А.С., Чорич П., Амазаспян А., Дэвис В., Аксман Р., Уайт МК, Абу-Гарбия М., Чайлдерс В., Кондра Дж. Х., Буазиз С., Сафак М. (октябрь 2016 г.). «Исходя из неизвестного: структурные и функциональные особенности агнопротеина полиомавирусов». Журнал клеточной физиологии. 231 (10): 2115–27. Дои:10.1002 / jcp.25329. ЧВК 5217748. PMID 26831433.
- ^ "Сайт таксономии ICTV".
- ^ Международное агентство по изучению рака (2013 г.). «Рабочая группа МАИР по оценке канцерогенного риска для людей. Малярия и некоторые полиомавирусы (вирусы SV40, BK, JC и клетки Меркеля)». Монографии МАИР по оценке канцерогенных рисков для человека. 104.
- ^ Перес-Лосада М., Кристенсен Р.Г., Макклеллан Д.А., Адамс Б.Дж., Вискиди Р.П., Демма Дж.С., Крэндалл К.А. (июнь 2006 г.). «Сравнение филогенетической дивергенции полиомавирусов и их хозяев». Журнал вирусологии. 80 (12): 5663–9. Дои:10.1128 / JVI.00056-06. ЧВК 1472594. PMID 16731904.
- ^ Джон Р., Бак CB, Алландер Т., Этвуд В.Дж., Гарси Р.Л., Империале М.Дж., майор Е.О., Рамквист Т., Норкин Л.К. (сентябрь 2011 г.). «Таксономические изменения в семействе Polyomaviridae». Архив вирусологии. 156 (9): 1627–34. Дои:10.1007 / s00705-011-1008-х. ЧВК 3815707. PMID 21562881.
- ^ Siqueira JD, Ng TF, Miller M, Li L, Deng X, Dodd E, Batac F, Delwart E (июль 2017 г.). «Эндемическая инфекция выброшенных на мель южных каланов (Enhydra lutris nereis) новым парвовирусом, полиомавирусом и аденовирусом». Журнал болезней дикой природы. 53 (3): 532–542. Дои:10.7589/2016-04-082. PMID 28192039.
- ^ Дела Круз Ф.Н., Ли Л., Делварт Э., Песавенто ПА (2017). «Новый полиомавирус легких у альпак (Vicugna pacos)». Ветеринарная микробиология. 201: 49–55. Дои:10.1016 / j.vetmic.2017.01.005. PMID 28284622.
- ^ Ци Д., Шань Т., Лю З., Дэн Х, Чжан З., Би В., Оуэнс Дж. Р., Фэн Ф, Чжэн Л., Хуан Ф, Делварт Э, Хоу Р., Чжан В. (2017). «Новый полиомавирус из носовой полости гигантской панды (Ailuropoda melanoleuca)». Журнал вирусологии. 14 (1): 207. Дои:10.1186 / s12985-017-0867-5. ЧВК 5658932. PMID 29078783.
- ^ Гонсалвес Мотта Майя Ф., Марсель де Соуза В., Сабино-Сантос Г., Хорхе Фумагалли М., Модха С., Рамиро Мурсия П., Тадеу Мораес Фигейредо Л. (2018). «Новый полиомавирус у сигмодонтиновых грызунов из штата Сан-Паулу, Бразилия». Архив вирусологии. 163 (10): 2913–2915. Дои:10.1007 / s00705-018-3913-8. PMID 29931397.
- ^ Лю П, Цю И, Син Ц, Чжоу Дж. Х., Ян У., Ван Ц, Ли Дж Й, Хан Х, Чжан И З, Ге XY (2019). «Обнаружение и характеристика генома двух новых вирусов папилломы и нового полиомавируса у землероек (Tupaia belangeri chinensis) в Китае». Вирол Дж. 16 (1): 35. Дои:10.1186 / s12985-019-1141-9. ЧВК 6423848. PMID 30885224.
- ^ Гейт Т., Датта С., Оливер Дж., Робитайл А., Хампрас С., Комб Дж. Д., Маккей-Шопен С., Ле Кальвез-Кельм Ф, Фенске Н., Черпелис Б., Джулиано А.Р., Франчески С., Маккей Дж., Роллисон Д.Е., Томмазино М ( 2017). «Выделение и характеристика нового предполагаемого полиомавируса человека». Вирусология. 506: 45–54. Дои:10.1016 / j.virol.2017.03.007. PMID 28342387.
- ^ Альтман Л.К. (18 января 2008 г.). «Вирус связан с сильным раком кожи». Нью-Йорк Таймс. Получено 18 января 2008.
- ^ Шуда М., Арора Р., Квун Х.Дж., Фенг Х., Сарид Р., Фернандес-Фигерас М.Т., Толстов Ю., Гьёруп О., Мансукхани М.М., Свердлов С.Х., Чаудхари П.М., Кирквуд Дж.М., Налесник М.А., Кант Я.А., Вайс Л.М., Мур П.С., Чанг И (сентябрь 2009 г.). "Человеческая полиомавирусная инфекция клеток Меркеля I. Экспрессия Т-антигена MCV в карциноме клеток Меркеля, лимфоидных тканях и лимфоидных опухолях". Международный журнал рака. 125 (6): 1243–9. Дои:10.1002 / ijc.24510. ЧВК 6388400. PMID 19499546.
- ^ ван дер Мейден Э, Янссенс Р. У., Лаубер С., Бувес Бавинк Дж. Н., Горбаленя А. Э., Feltkamp MC (июль 2010 г.). «Открытие нового полиомавируса человека, связанного со спинулезной триходисплазией, у пациента с ослабленным иммунитетом». Патогены PLOS. 6 (7): e1001024. Дои:10.1371 / journal.ppat.1001024. ЧВК 2912394. PMID 20686659.
- ^ Kazem S, van der Meijden E, Feltkamp MC (август 2013 г.). «Полиомавирус, связанный с триходисплазией spinulosa: вирусологические предпосылки и клинические последствия». APMIS. 121 (8): 770–82. Дои:10.1111 / apm.12092. PMID 23593936.
- ^ Скада Н., Хофманн Дж., Кальвиньяк-Спенсер С., Рупрехт К., Лиман П., Кюн Дж., Хенгель Х., Элерс Б. (май 2011 г.). «Новый полиомавирус человека, тесно связанный с лимфотропным полиомавирусом африканской зеленой обезьяны». Журнал вирусологии. 85 (9): 4586–90. Дои:10.1128 / jvi.02602-10. ЧВК 3126223. PMID 21307194.
- ^ Korup S, Rietscher J, Calvignac-Spencer S, Trusch F, Hofmann J, Moens U, Sauer I, Voigt S, Schmuck R, Ehlers B (2013). «Выявление нового полиомавируса человека в органах желудочно-кишечного тракта». PLOS One. 8 (3): e58021. Bibcode:2013PLoSO ... 858021K. Дои:10.1371 / journal.pone.0058021. ЧВК 3596337. PMID 23516426.
- ^ Мишра Н., Перейра М., Родос Р. Х., Ан П, Пипас Дж. М., Джайн К., Капур А., Бризе Т., Фауст П.Л., Липкин В.И. (ноябрь 2014 г.). «Идентификация нового полиомавируса у реципиента трансплантата поджелудочной железы со слепотой сетчатки и васкулитной миопатией». Журнал инфекционных болезней. 210 (10): 1595–9. Дои:10.1093 / infdis / jiu250. ЧВК 4334791. PMID 24795478.
- ^ Гарднер С.Д., Филд А.М., Коулман Д.В., Халм Б. (июнь 1971 г.). «Новый паповавирус человека (B.K.), выделенный из мочи после трансплантации почки». Ланцет. 1 (7712): 1253–7. Дои:10.1016 / с0140-6736 (71) 91776-4. PMID 4104714.
- ^ Padgett BL, Walker DL, ZuRhein GM, Eckroade RJ, Dessel BH (июнь 1971 г.). «Культивирование паповируса из головного мозга человека с прогрессирующей мультифокальной лейкоэнцефалопатией». Ланцет. 1 (7712): 1257–60. Дои:10.1016 / S0140-6736 (71) 91777-6. PMID 4104715.
- ^ Алландер Т., Андреассон К., Гупта С., Бьеркнер А., Богданович Г., Перссон М.А., Далианис Т., Рамквист Т., Андерссон Б. (апрель 2007 г.). «Выявление третьего полиомавируса человека». Журнал вирусологии. 81 (8): 4130–6. Дои:10.1128 / JVI.00028-07. ЧВК 1866148. PMID 17287263.
- ^ а б Nguyen KD, Lee EE, Yue Y, Stork J, Pock L, North JP, Vandergriff T, Cockerell C, Hosler GA, Pastrana DV, Buck CB, Wang RC (май 2017 г.). «Полиомавирусы человека 6 и 7 связаны с зудящими и дискератотическими дерматозами». Журнал Американской академии дерматологии. 76 (5): 932–940.e3. Дои:10.1016 / j.jaad.2016.11.035. ЧВК 5392424. PMID 28040372.
- ^ Хо Дж, Джедрих Дж. Дж., Фен Х, Натали А. А., Грандинетти Л., Мирвиш Е., Креспо М. М., Ядав Д., Фасанелла К. Э., Прокселл С., Куан С.Ф., Пастрана Д.В., Бак CB, Шуда Ю., Мур П.С., Чанг И (май 2015 г.) ). «Связанная с полиомавирусом 7 человека зудящая сыпь и виремия у реципиентов трансплантата». Журнал инфекционных болезней. 211 (10): 1560–5. Дои:10.1093 / infdis / jiu524. ЧВК 4425822. PMID 25231015.
- ^ Toptan T, Yousem SA, Ho J, Matsushima Y, Stabile LP, Fernández-Figueras MT, Bhargava R, Ryo A, Moore PS, Chang Y (февраль 2016 г.). «Исследование полиомавирусов человека при раке». JCI Insight. 1 (2). Дои:10.1172 / jci.insight.85562. ЧВК 4811373. PMID 27034991.
- ^ Siebrasse EA, Reyes A, Lim ES, Zhao G, Mkakosya RS, Manary MJ, Gordon JI, Wang D (октябрь 2012 г.). «Идентификация полиомавируса MW, нового полиомавируса в стуле человека». Журнал вирусологии. 86 (19): 10321–6. Дои:10.1128 / JVI.01210-12. ЧВК 3457274. PMID 22740408.
- ^ Бак CB, Phan GQ, Raiji MT, Murphy PM, McDermott DH, McBride AA (октябрь 2012 г.). «Полная последовательность генома десятого полиомавируса человека». Журнал вирусологии. 86 (19): 10887. Дои:10.1128 / JVI.01690-12. ЧВК 3457262. PMID 22966183.
- ^ Ю. Г., Гренингер А. Л., Иса П., Фан Т. Г., Мартинес М. А., де ла Лус Санчес М., Контрерас Дж. Ф., Сантос-Пресьядо Дж. И., Парсоннет Дж., Миллер С., ДеРизи Дж. Л., Делварт Е., Ариас К. Ф., Чиу С. Ю. (2012). «Открытие нового полиомавируса в образцах острой диареи у детей». PLOS One. 7 (11): e49449. Bibcode:2012PLoSO ... 749449Y. Дои:10.1371 / journal.pone.0049449. ЧВК 3498111. PMID 23166671.
- ^ Лим Э.С., Рейес А., Антонио М., Саха Д., Икумапайи ООН, Адейеми М., Стайн О.К., Скелтон Р., Бреннан, округ Колумбия, Мкакося Р.С., Манари М.Дж., Гордон Д.И., Ван Д. (февраль 2013 г.). «Открытие полиомавируса STL, полиомавируса предкового рекомбинантного происхождения, который кодирует уникальный Т-антиген путем альтернативного сплайсинга». Вирусология. 436 (2): 295–303. Дои:10.1016 / j.virol.2012.12.005. ЧВК 3693558. PMID 23276405.
- ^ Гейт, Тарик; Датта, Санкхадип; Оливер, Хавьер; Робитайл, Алексис; Хампрас, Шалака; Комб, Жан-Дамьен; Маккей-Шопен, Сандрин; Кальвез-Кельм, Флоренс Ле; Фенске, Нил (2017). «Выделение и характеристика нового предполагаемого полиомавируса человека». Вирусология. 506: 45–54. Дои:10.1016 / j.virol.2017.03.007. PMID 28342387.
- ^ Prado JC, Monezi TA, Amorim AT, Lino V, Paladino A, Boccardo E (2018). «Полиомавирусы человека и рак: обзор». Клиники (Сан-Паулу). 73 (приложение 1): e558s. Дои:10.6061 / клиники / 2018 / e558s. ЧВК 6157077. PMID 30328951.
- ^ Далианис Т., Хирш Х.Х. (март 2013 г.). «Полиомавирусы человека при болезнях и раках». Вирусология. 437 (2): 63–72. Дои:10.1016 / j.virol.2012.12.015. PMID 23357733.
- ^ Ван Гелу М., Хан М.Т., Элерс Б., Моэнс У. (ноябрь 2012 г.). «Геномный анализ новых полиомавирусов человека». Обзоры в медицинской вирусологии. 22 (6): 354–77. Дои:10.1002 / rmv.1711. PMID 22461085.
- ^ Эгли А., Инфанти Л., Думулин А., Базер А., Самаридис Дж., Стеблер С., Госерт Р., Хирш Х. Х. (март 2009 г.). «Распространенность и репликация полиомавирусов BK и JC у 400 здоровых доноров крови». Журнал инфекционных болезней. 199 (6): 837–46. Дои:10.1086/597126. PMID 19434930.
- ^ а б Poulin DL, DeCaprio JA (сентябрь 2006 г.). «Есть ли роль SV40 в раке человека?». Журнал клинической онкологии. 24 (26): 4356–65. Дои:10.1200 / JCO.2005.03.7101. PMID 16963733.
- ^ а б zur Hausen H (декабрь 2003 г.). «SV40 при раке человека - бесконечная сказка?». Международный журнал рака. 107 (5): 687. Дои:10.1002 / ijc.11517. PMID 14566815.
- ^ Gazdar AF, Butel JS, Carbone M (декабрь 2002 г.). «SV40 и опухоли человека: миф, связь или причинная связь?». Обзоры природы. Рак. 2 (12): 957–64. Дои:10.1038 / nrc947. PMID 12459734.
- ^ Кэрролл-Панкхерст С., Энгельс Е.А., Стриклер HD, Годерт Дж. Дж., Вагнер Дж., Мортимер Е. А. (ноябрь 2001 г.). «Тридцатипятилетняя смертность после получения вакцины против полиомиелита, зараженной SV40, в неонатальный период». Британский журнал рака. 85 (9): 1295–7. Дои:10.1054 / bjoc.2001.2065. ЧВК 2375249. PMID 11720463.
- ^ Шах К.В. (январь 2007 г.). «SV40 и рак человека: обзор последних данных». Международный журнал рака. 120 (2): 215–23. Дои:10.1002 / ijc.22425. PMID 17131333.
- ^ а б Drachenberg CB, Hirsch HH, Ramos E, Papadimitriou JC (декабрь 2005 г.). «Полиомавирусная болезнь при трансплантации почки: обзор патологических данных и методов диагностики». Патология человека. 36 (12): 1245–55. Дои:10.1016 / j.humpath.2005.08.009. PMID 16311117.
- ^ Viscidi RP, Clayman B (2006). «Серологическая перекрестная реактивность между капсидами полиомавируса». В Ахсане Н (ред.). Полиомавирусы и болезни человека. Успехи экспериментальной медицины и биологии. 577. С. 73–84. Дои:10.1007/0-387-32957-9_5. ISBN 978-0-387-29233-5. PMID 16626028.
- ^ Дрюс К., Башир Т., Дёррис К. (январь 2000 г.). «Количественное определение полиомавируса человека JC в ткани мозга и спинномозговой жидкости пациентов с прогрессирующей мультифокальной лейкоэнцефалопатией с помощью конкурентной ПЦР». Журнал вирусологических методов. 84 (1): 23–36. Дои:10.1016 / S0166-0934 (99) 00128-7. PMID 10644084.
- ^ Никелейт В., Хирш Х. Х., Бине И. Ф., Гудат Ф., Принц О., Далкен П., Тиль Г., Михатч М. Дж. (Май 1999 г.). «Полиомавирусная инфекция реципиентов почечного аллотрансплантата: от латентной инфекции до манифестного заболевания». Журнал Американского общества нефрологов. 10 (5): 1080–9. PMID 10232695.
- ^ Randhawa PS, Vats A, Zygmunt D, Swalsky P, Scantlebury V, Shapiro R, Finkelstein S (август 2002). «Количественное определение вирусной ДНК в ткани почечного аллотрансплантата от пациентов с вирусной нефропатией BK». Трансплантация. 74 (4): 485–8. Дои:10.1097/00007890-200208270-00009. PMID 12352906.
- ^ Busam KJ, Jungbluth AA, Rekthman N, Coit D, Pulitzer M, Bini J, Arora R, Hanson NC, Tassello JA, Frosina D, Moore P, Chang Y (сентябрь 2009 г.). «Экспрессия полиомавируса клеток Меркеля в карциномах клеток Меркеля и ее отсутствие в комбинированных опухолях и нейроэндокринных карциномах легких». Американский журнал хирургической патологии. 33 (9): 1378–85. Дои:10.1097 / PAS.0b013e3181aa30a5. ЧВК 2932664. PMID 19609205.
- ^ Толстов Ю.Л., Пастрана Д.В., Фенг Х., Беккер Дж.К., Дженкинс Ф.Дж., Moschos S, Чанг Й., Бак CB, Мур PS (сентябрь 2009 г.). «Человеческая полиомавирусная инфекция клеток Меркеля II. MCV - это обычная человеческая инфекция, которая может быть обнаружена с помощью иммуноанализа конформационных капсидных эпитопов». Международный журнал рака. 125 (6): 1250–6. Дои:10.1002 / ijc.24509. ЧВК 2747737. PMID 19499548.
- ^ Пастрана Д.В., Толстов Ю.Л., Беккер Дж. К., Мур П.С., Чанг Й., Бак CB (сентябрь 2009 г.). «Количественное определение серореактивности человека к полиомавирусу клеток Меркеля». Патогены PLOS. 5 (9): e1000578. Дои:10.1371 / journal.ppat.1000578. ЧВК 2734180. PMID 19750217.
- ^ Картер Дж. Дж., Полсон К. Г., Wipf GC, Миранда Д., Мадлен М. М., Джонсон Л. Г., Лемос Б. Д., Ли С., Варкола А. Х., Айер Дж. Г., Нгием П., Галлоуэй Д. А. (ноябрь 2009 г.). «Ассоциация антител, специфичных к полиомавирусу клеток Меркеля, с карциномой из клеток Меркеля». Журнал Национального института рака. 101 (21): 1510–22. Дои:10.1093 / jnci / djp332. ЧВК 2773184. PMID 19776382.
- ^ Элизабет Матисоо-Смит; К. Энн Хорсбург (2012). ДНК для археологов. Рутледж. ISBN 978-1598746815.
- ^ Гросс L (июнь 1953 г.). «Фильтруемый агент, извлеченный из лейкозных экстрактов Ak, вызывающий карциному слюнных желез у мышей C3H». Труды Общества экспериментальной биологии и медицины. 83 (2): 414–21. Дои:10.3181/00379727-83-20376. PMID 13064287.
- ^ Стюарт С.Е., Эдди Б.Е., Боргезе Н. (июнь 1958 г.). «Новообразования у мышей, инокулированных опухолевым агентом, внесенным в культуру ткани». Журнал Национального института рака. 20 (6): 1223–43. Дои:10.1093 / jnci / 20.6.1223. PMID 13549981.
- ^ Эдди Б. Е., Стюарт С. Е. (ноябрь 1959 г.). «Характеристики вируса полиомы SE». Американский журнал общественного здравоохранения и здоровья нации. 49 (11): 1486–92. Дои:10.2105 / AJPH.49.11.1486. ЧВК 1373056. PMID 13819251.
- ^ Шовальтер Р.М., Пастрана Д.В., Бак CB (июль 2011 г.). «Гликозаминогликаны и сиалированные гликаны последовательно способствуют проникновению инфекции полиомавируса клеток Меркеля». Патогены PLOS. 7 (7): e1002161. Дои:10.1371 / journal.ppat.1002161. ЧВК 3145800. PMID 21829355.
- ^ Готтлиб К.А., Вильярреал LP (июнь 2001 г.). «Естественная биология среднего Т-антигена полиомавируса». Обзоры микробиологии и молекулярной биологии. 65 (2): 288–318, вторая и третья страницы, оглавление. Дои:10.1128 / mmbr.65.2.288-318.2001. ЧВК 99028. PMID 11381103.
- ^ Перетти А., Фитцджеральд ПК, Блисковский В., Пастрана Д.В., Бак CB (январь 2015 г.). «Последовательность генома полиомавируса, ассоциированного с рыбами, полиомавируса 1 черноморского окуня (Centropristis striata)». Анонсы генома. 3 (1): e01476-14. Дои:10.1128 / genomeA.01476-14. ЧВК 4319505. PMID 25635011.
- ^ Лопес-Буэно А., Мавиан С., Лабелла А. М., Кастро Д., Боррего Дж. Дж., Альками А., Алехо А. (октябрь 2016 г.). «Встречаемость иридовируса, полиомавируса и уникального члена новой группы папилломавирусов рыб у морского леща, пораженного лимфоцистозом». Журнал вирусологии. 90 (19): 8768–79. Дои:10.1128 / JVI.01369-16. ЧВК 5021401. PMID 27440877.